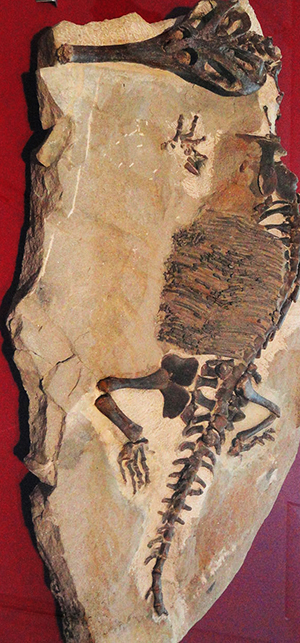
Champsosaurus annectens Cope, 1876
(Da: it.wikipedia.org)
Phylum: Chordata Haeckel, 1874
Subphylum: Vertebrata Cuvier, 1812
Classe: Sauropsida Huxley, 1864
Ordine: Choristodera Cope, 1884
Famiglia: Champsosauridae Cope, 1884
Genere: Champsosaurus Cope, 1877
Descrizione
Lunghi in media fra il metro e i tre metri, questi animali erano piuttosto simili a piccoli coccodrilli, con un corpo lungo e appiattito, una lunga coda, arti piuttosto brevi e un cranio allungato. Le fauci erano munite di molti denti aguzzi, ed erano perfette per catturare i pesci dei quali i coristoderi si nutrivano. Il cranio presenta occhi posti molto in avanti, un'estremità posteriore molto allargata, ospitante grandi muscoli che azionavano le fauci, e un muso lunghissimo simile a quello dell'attuale gaviale del Gange (Gavialis gangeticus Gmelin J.F., 1789). A causa di queste somiglianze, i paleontologi ritengono che questi rettili conducessero uno stile di vita paragonabile a quello dei coccodrilli. I coristoderi più primitivi, in ogni caso, possedevano crani più corti e larghi. Probabilmente i coristoderi cacciavano piccoli animali nelle paludi e negli specchi d'acqua dolce, muovendo la lunga coda appiattita lateralmente per spostarsi, e direzionando il corpo grazie alle zampe appiattite e palmate. Il primo fossile di un coristodero fu rinvenuto nel 1876 in Montana, in strati del Cretaceo superiore, e venne descritto da Edward Drinker Cope come Champsosaurus, ovvero "lucertola coccodrillo". Da allora sono stati rinvenuti molti altri fossili di questi animali (in particolare in strati che vanno dal tardo Cretaceo all'Eocene inferiore - da 75 a 50 milioni di anni fa), ma la loro classificazione è tutt'altro che chiara. Alcuni studiosi li ritengono stretti parenti di lucertole e serpenti, altri pensano che i coristoderi fossero arcosauromorfi primitivi. In ogni caso, questi animali erano rettili diapsidi, ovvero muniti di due "finestre" per ogni lato del cranio, per ospitare i muscoli delle mascelle. L'evoluzione dei coristoderi ebbe inizio da un antenato diapside sconosciuto, probabilmente già nel Triassico superiore, circa 220 milioni di anni fa. Resti appartenenti a un animale denominato Pachystropheus rhaeticus e forse ascrivibile al gruppo, risalgono appunto a quel periodo. La mancanza di materiale cranico, però, induce alla cautela i paleontologi. Nel Giurassico medio apparvero i primi veri coristoderi: Monjurosuchus della Cina e Cteniogenys, vissuto in Nordamerica ed Europa (in passato considerato un parente delle "lucertole volanti" Kuehneosaurus e Icarosaurus), sono esempi di piccoli coristoderi primitivi. Altri due coristoderi del Cretaceo inferiore, Shokawa del Giappone e Hyphalosaurus della Cina, possiedono strani colli allungati; in questo assomigliavano ai plesiosauri e ai notosauri. Altri coristoderi più evoluti, appartenenti alle famiglie Champsosauridae e Simoedosauridae, sono raggruppati nei Neochoristodera, che include forme quali Champsosaurus, Simoedosaurus, Tchoiria e Ikechosaurus. Questo gruppo comprende i coristoderi più grandi e maggiormente adattati al nuoto. Diversamente da molti altri gruppi di rettili, questi animali sopravvissero all'estinzione del Cretaceo-Terziario: i loro fossili sono infatti comuni in sedimenti risalenti al Paleocene e all'Eocene (circa 50 milioni di anni fa). Alcuni campsosauridi siano stati rinvenuti in Alaska, in terreni del Cretaceo superiore: ciò indica che, a quell'epoca, il clima di quei luoghi era molto più mite. In media, i campsosauri del Cretaceo raggiungevano la lunghezza di un metro, ma una forma del Paleocene, Champsosaurus gigas, era lunga fino a 3 metri. Fino agli anni novanta si pensava che i coristoderi si fossero estinti nell'Eocene. La scoperta, avvenuta nel 1992, di un animale denominato Lazarussuchus inexpectatus ("coccodrillo Lazzaro inaspettato"), ha esteso di undici milioni di anni la distribuzione stratigrafica del gruppo (tardo Oligocene). Successive analisi cladistiche hanno poi portato a ipotizzare che Lazarussuchus fosse molto più primitivo dei suoi parenti eocenici e addirittura più primitivo del presunto coristodero triassico Pachystopheus. Si ipotizza che Lazarussuchus potesse far parte di una linea fantasma originatasi almeno nel Cretaceo. Altri resti di questo animale sono poi stati scoperti in strati del Miocene inferiore (circa 20 milioni di anni fa). I fossili sono stati rinvenuti in Francia e in Repubblica Ceca. L'estinzione dei coristoderi potrebbe essere stata causata da una serie di fattori, tra i quali l'abbassamento della temperatura nel corso dell'Oligocene, l'aumento della competizione da parte dei coccodrilli, la perdita di habitat, o altre combinazioni di fattori. Nel 2006, la Royal Society del Regno Unito annunciò la scoperta di un coristodero fossile munito di due teste; questa fu la prima volta di un ritrovamento di questo tipo. Le possibilità di ritrovare un fossile simile sono molto basse, come dimostra la proporzione corrente di rettili nati con due teste.
Diffusione
Vissuti tra il Giurassico e il Miocene (tra 160 e 20 milioni di anni fa). I loro resti sono stati rinvenuti in Nordamerica, Asia ed Europa.
Bibliografia
Cope ED (1876). "On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana". Proceedings of the Academy of Natural Sciences of Philadelphia. 1876: 340-359.
Dudgeon, Thomas William (2019). The internal cranial anatomy of Champsosaurus lindoei and its functional implications (PDF). Earth Sciences (M.Sci.). Ottawa, Ontario: Carleton University.
B. Brown. 1905. The osteology of Champsosaurus Cope. American Museum of Natural History, Memoirs 9:1-264.
W. A. Parks. 1933. New species of Champsosaurus from the Belly River Formation of Alberta, Canada. Transactions of Royal Society of Canada 27:121-137.
W. A. Parks. 1927. Champsosaurus albertensis, a new species of rhynchocephalian from the Edmonton Formation of Alberta. University of Toronto Studies of Geological Survey 25:1-48.
Sigogneau-Russell D. 1979. Les champsosaures Européens: mise au point sur le champsosaure d'erquelinnes (landenien inferieur, Belgique). Annales de Paliontologit (Vertibris) 65: 7-62.
B. R. Erickson. 1972. The lepidosaurian reptile Champsosaurus in North America. The Science Museum of Minnesota, Monograph (Paleontology) 1:1-91.
Gao, Keqin; Fox, Richard C. (December 1998). "New choristoderes (Reptilia: Diapsida) from the Upper Cretaceous and Palaeocene, Alberta and Saskatchewan, Canada, and phylogenetic relationships of Choristodera". Zoological Journal of the Linnean Society. 124 (4): 303-353.
Lehman, Thomas M.; Barnes, Ken (2010). "Champsosaurus (Diapsida: Choristodera) from the Paleocene of west Texas: paleoclimatic implications". Journal of Paleontology. 84 (2): 341-345.
Vandermark, Deborah; Tarduno, John A.; Brinkman, Donald B. (May 2007). "A fossil champsosaur population from the high Arctic: Implications for Late Cretaceous paleotemperatures". Palaeogeography, Palaeoclimatology, Palaeoecology. 248 (1-2): 49-59.
D.Lambert, D.Naish and E.Wyse 2001, "Encyclopedia of Dinosaurs and prehistoric life", p. 77, Dorling Kindersley Limited, London.
Hoganson, J.W.; J. Campbell (1995). "Restoration and display of a crocodilie-like Champsosaurus gigas skeleton at the North Dakota Heritage Center" (PDF). NDGS Newsletter. 22 (3): 8-10.
"Champsosaurus gigas" (PDF). Department of Mineral Resources, North Dakota.
Erickson BR (June 1985). "Aspects of some anatomical structures of Champsosaurus (Reptilia: Eosuchia)". Journal of Vertebrate Paleontology. 5 (2): 111-127.
Dudgeon, Thomas W.; Maddin, Hillary C.; Evans, David C.; Mallon, Jordan C. (April 2020). "Computed tomography analysis of the cranium of Champsosaurus lindoei and implications for the choristoderan neomorphic ossification". Journal of Anatomy. 236 (4): 630-659.

|
Data: 01/12/2011
Emissione: Coccodrilli preistorici Stato: Burundi Nota: Emesso in un foglietto di 4 v. diversi |
|---|

|
Data: 01/12/2011
Emissione: Coccodrilli preistorici Stato: Burundi |
|---|